CURRENT RESEARCH PROJECTS OF THE POLLMANN LAB - Auxin metabolism
Indole-3-acetic acid (IAA) is the major naturally occurring auxin. Over the past decades, evidence has accumulated both emphasising the physiological importance of auxin in the context of coordinating plant development and describing the molecular mode of auxin action. However, although IAA constitutes a rather simple molecule, sharing major structural features with the proteinogenic amino acid L-tryptophan, the biosynthesis of IAA remained elusive for a very long time.
IAA is one of the major growth factors in plants. It is recognized to be involved in virtually all aspects of plant growth and development.
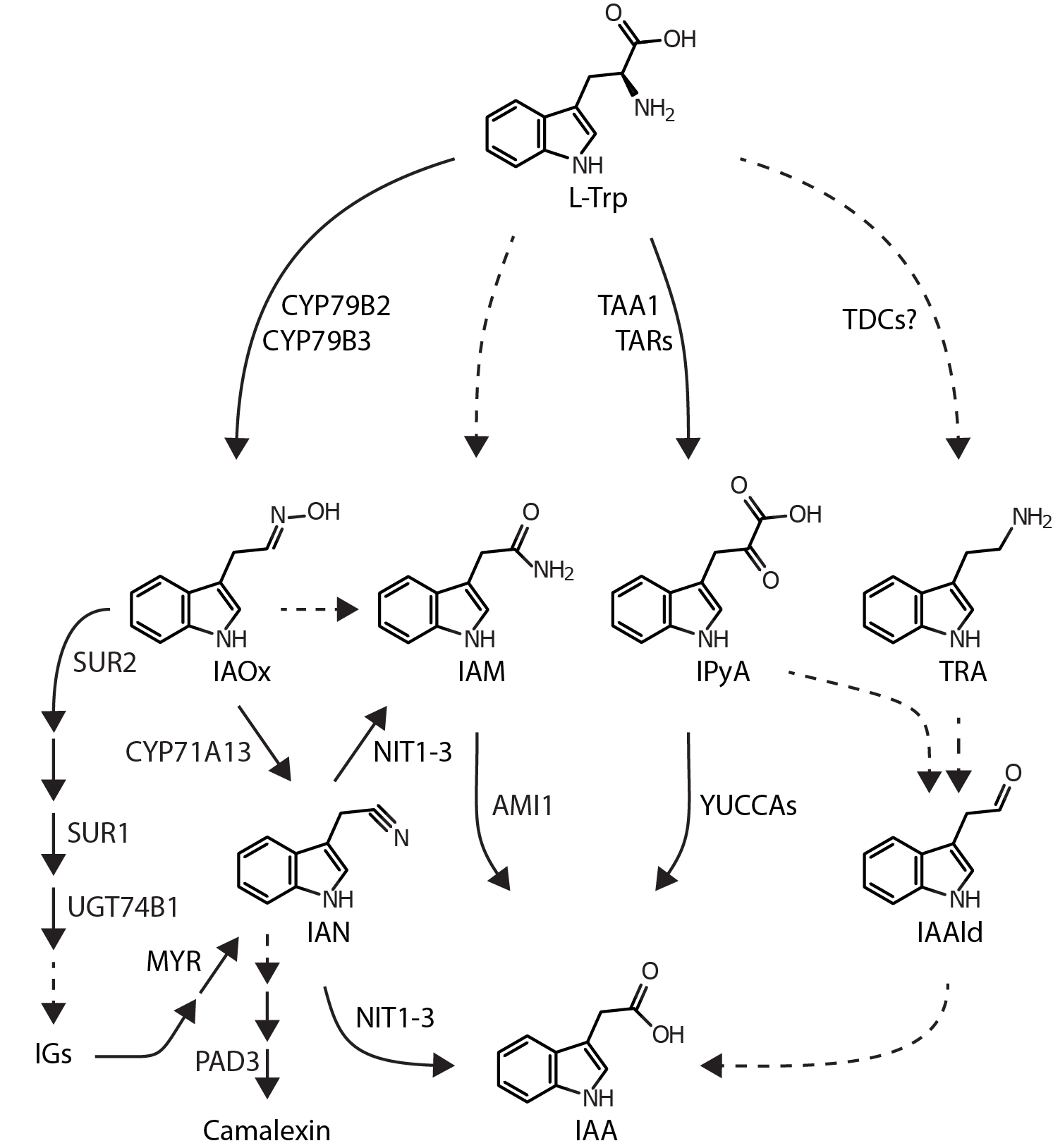
In recent years, experimental proof has been provided that auxin biosynthesis in plants is realised by a small number of alternative pathways, each of them designated for an intermediate that is a hallmark of the pathway (Fig. 1).
Figure 1. Proposed anabolic routes for IAA biosynthesis in Arabidopsis thaliana. The IAOx pathway that is seemingly restricted to IG-producing plant species is given on the left. The IPyA pathway via TAR1/TAR2 and the YUCCA enzymes is the major route for auxin biosynthesis. Dashed lines represent assumed reaction steps for which the corresponding genes/enzymes have not yet been identified. Metabolites and enzymes are abbreviated as follows: AMI1, amidase 1; CYP71A13, cytochrome P450 monooxygenase 71A13; CYP79B2/B3, cytochrome P450 monooxygenase 79B2/B3; IAA, indole-3-acetic acid; IAAld, indole-3-acetaldehyde; IAM, indole-3-acetamide; IAN, indole-3-acetonitrile; IAOx, indole-3-acetaldoxime; IGs, indole glucosinolates; IPyA, indole-3-pyruvic acid; l-Trp, l-tryptophan; MYR, myrosinase; NIT1-3, nitrilase 1-3; PAD3 PHYTOALEXIN DEFICIENT 3 (cytochrome P450 monooxygenase 71B15); SUR1, SUPERROOT 1 (S-alkyl-thiohydroxymate lyase); SUR2, SUPERROOT 2 (cytochrome P450 monooxygenase 83B1); TAA1, tryptophan aminotransferase of Arabidopsis 1; TAR, tryptophan aminotransferase related; TDC, tryptophan decarboxylase; TRA, tryptamine; UGT74B1, UDP-glucosyl transferase 74B1.
Thus far, four Trp-dependent pathways for auxin biosynthesis have been proposed. These are the indole-3-acetaldoxime (IAOx)-pathway, the tryptamine (TAM)-pathway, the indole-3-pyruvic acid (IPyA)-pathway, and the indole-3-acetamide (IAM)-pathway. Some people also consider a Trp-independent pathway for the formation of IAA, but genetic and molecular evidence for its existence is, so far, missing. As yet, only one of the proposed pathways, the IPyA route, is fully disclosed with respect to the catalysed reaction steps and the enzymes involved. Due to the obvious gaps in the pathways, the functional redundancy, and the tissue and plant specific variations in expression patterns of the identified components, the relevance of each of these pathways is difficult to assess.
One major research line of our lab concerns auxin biosynthesis and its regulation in the model plant Arabidopsis thaliana. Our previous work provided evidence that led us to suggest that one route of auxin formation takes its course via the intermediate IAM, a compound proven to be endogenous to Arabidopsis and several other plant species (Pollmann et al. 2002; Sugawara et al. 2009). In the past, we have succeeded in identifying and characterising the first plant IAM hydrolase (AMI1) from Arabidopsis, capable of catalysing the conversion of IAM to IAA (for review, see: Pollmann et al. 2006; Lehmann et al. 2010). AMI1 is located in the cytoplasm (Pollmann et al. 2006), the assumed main locus of IAA biosynthesis. Judging by its primary amino acid composition and homology to other well-characterised enzymes, AMI1 is considered a member of the amidase signature family that comprises enzymes that are found widespread in nature, catalysing a diverse range of different reactions. To date, more than 20 AMI1-like proteins have been identified from monocot and dicot plant species (Mano et al. 2010; Lehmann et al. 2010, Sánchez-Parra et al. 2014), suggesting a conserved and likely important function of AMI1-like enzymes. Our recent work on the elucidation of the role of AMI1-mediated IAA formation and on how AMI1 integrates into the already deciphered framework of auxin biosynthesis in plants revealed some intriguing aspects on a novel role of IAM as a negative plant growth regulator (Fig. 2).
Figure 2. Characterisation of ami1 loss-of-function mutants. Phenotype of Arabidopsis wild-type (Col-0) and ami1-2 plants grown for either 2 weeks on 0.5x MS plates or 4 weeks on soil. The IAM accumulating ami1 knockout mutants show a moderately reduced growth when grown on soil without additional carbon sources (Pérez-Alonso et al. 2021).
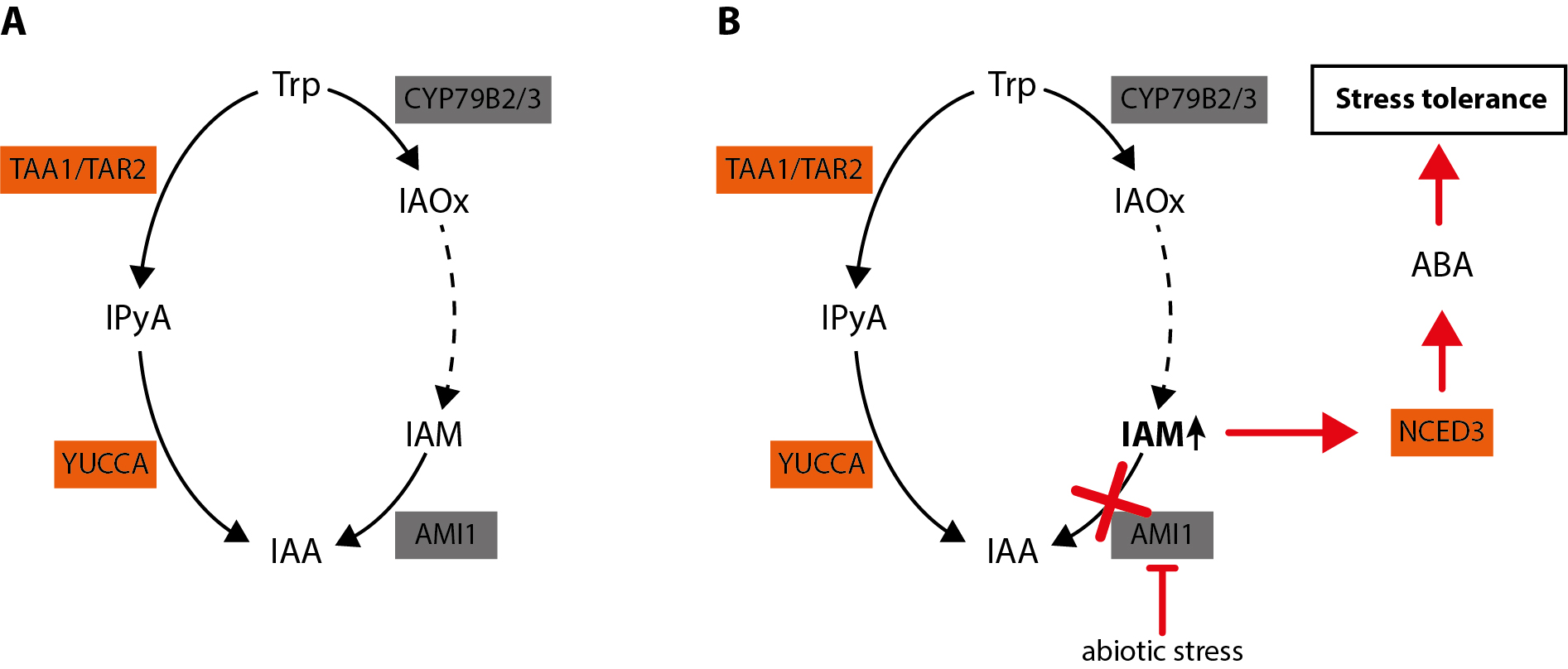
The accumulation of IAM has a growth reducing effect and triggers the crosstalk with other plant hormones, including abscisic acid and jasmonic acid
Figure 3. Integrated model of the molecular role of AMI1. (A) Our data suggest that the metabolic flux of the parallel pathways normally leads without distortions to IAA. In this scenario, IAM is not considerably accumulating as it is quickly converted into IAA. (B) Under stress conditions, however, the expression of AMI1 is repressed. In consequence, this results in the accumulation of IAM. The increasing IAM levels gradually induce the expression of NCED3 and trigger the subsequent biosynthesis of ABA as well as the corresponding downstream processes that confer stress resistance in Arabidopsis (Pérez-Alonso et al. 2021).
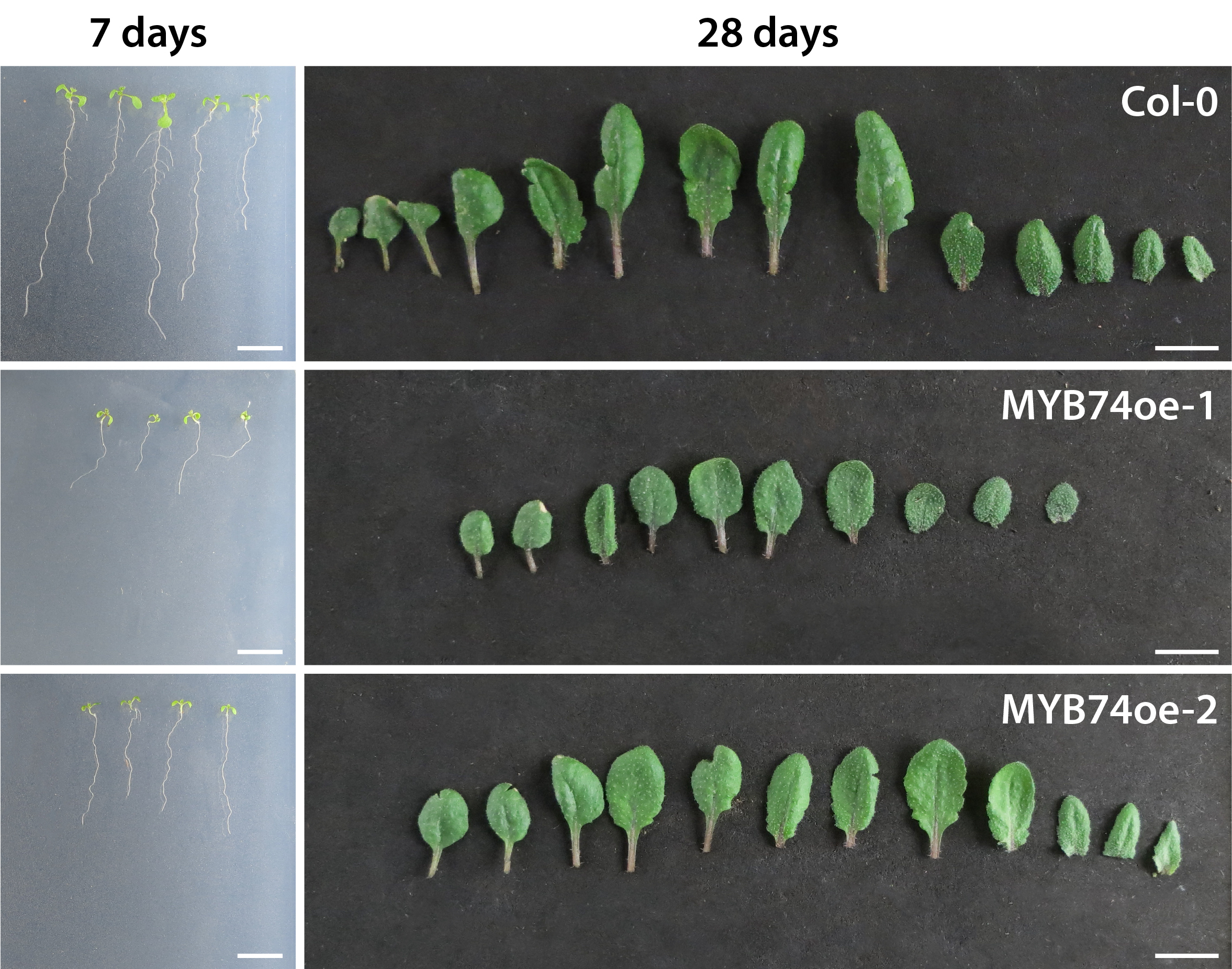
Just recently, we provided evidence that IAM and IAA transcriptionally regulate distinct subgroups of target genes (Ortiz-García et al. 2022). Among the IAM-responsive target genes, we identified and characterised the myeloblastosis (MYB) transcription factor MYB74. Our work reverse genetics, phenotypic, and transcriptomics analysis of the factor disclosed MYB74 as negative plant growth regulator (Fig. 4) and associated it with with abiotic stress responses, especially osmotic stress, in Arabidopsis.
Figure 4. Phenotypic analysis of conditional MYB74 overexpressor plants. Phenotypic appearance of wild-type (Col-0), MYB74oe-1, and MYB74oe-2 plants grown either for seven days on sterile 0.5X MS plates (left) or for 28 days on soil (right). Scale bars = 1 cm.